Archives
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-07
-
br Molecular genetics Based on
2020-05-11
Molecular genetics Based on the genetic mutations associated with their development, sarcomas are subdivided into two distinct classes. One class is composed of tumors bearing complex karyotypic abnormalities with no particular pattern. The second class, which includes Ewing sarcoma, encompasses
-
An Emerging Role for the EphrinB EphB RASA Signaling Axis
2020-05-11
An Emerging Role for the EphrinB2-EphB4-RASA1 Signaling Axis in Human Congenital Cerebrovascular Disorders Normal vascular development includes de novo blood vessel formation from endothelial cells (ECs) (vasculogenesis) 1, 2, expansion of the network through vessel sprouting, splitting, and remod
-
br Introduction Hydrogen sulfide H S and
2020-05-11
Introduction Hydrogen sulfide (H2S) and nitric oxide (NO) are important gasotransmitters in the cardiovascular system and instrumental to the fine control of vascular tone [1] and cellular function [2]. NO is synthesized from l-arginine by calmodulin-dependent endothelial nitric oxide synthase (e
-
Introduction Pain is frequently the first clinical symptom o
2020-05-11
Introduction Pain is frequently the first clinical symptom of head and neck carcinoma. Indeed, pain is the initial reason leading ∼70% of cancer patients to seek professional assistance (Lam & Schmidt, 2011; Rettig & D’Souza, 2015; Schmidt, 2014). Clinically, cancer-induced facial pain can be evoke
-
Abbas et al studied the mitigation of ASR in their
2020-05-09
Abbas et al. studied the mitigation of ASR in their corresponding study by incorporating RHA [1]. These researchers used Ordinary Portland Cement (OPC), four different percentages (i.e., 10%, 20%, 30% and 40% by weight of OPC) of RHA and reactive aggregate (sand from Dolomite-limestone rock) in thei
-
Oysters belong to the second largest animal phylum Mollusca
2020-05-09
Oysters belong to the second largest animal phylum Mollusca, and they are globally distributed aquaculture animals (Wang et al., 2018). A number of studies have demonstrated that oysters have evolved an integrated and complex innate immune system to recognize and eliminate various invaders. An array
-
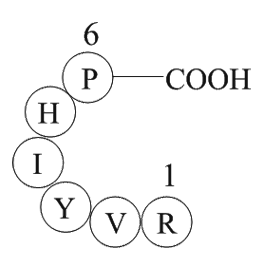
The murine subfamily members including
2020-05-09
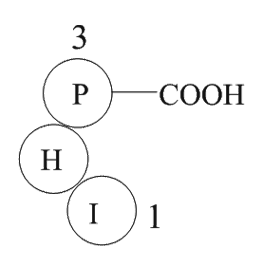
The murine subfamily members including , , , and have been identified and their encoded proteins showed deubiquitinating enzymatic activity , , , . It has been demonstrated that is an immediate-early gene induced by IL-3 in B-lymphocytes and is an IL-2 specific immediate-early gene in T-lymphoc
-
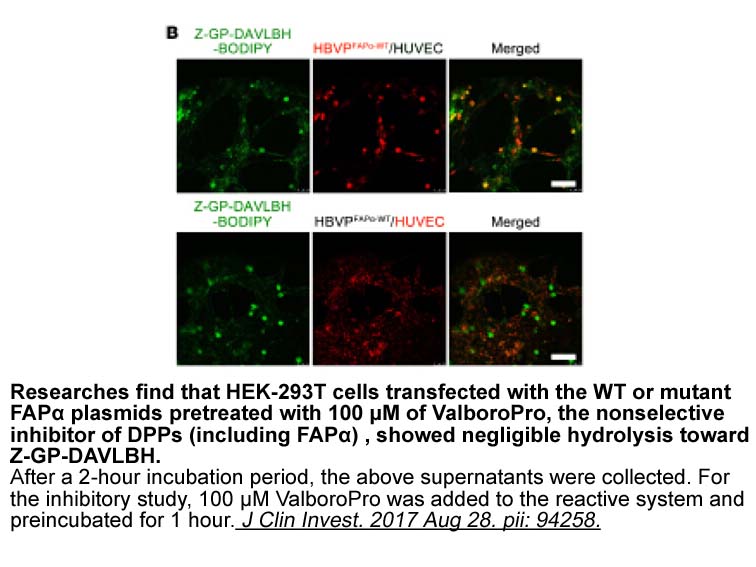
Fluorescein RNA Alpha SYN has no direct effect on forskolin
2020-05-09

Alpha-SYN has no direct effect on forskolin-induced CREB phosphorylation, but rather blocks the stimulation of CRE-mediated transcription in the nucleus. Previously we suggested that α-SYN can enter the nucleus and is present in both cytosolic and nuclear fractions without direct interaction between
-
The Lysine mutant is located in the tail region
2020-05-09
The Lysine429 mutant is located in the tail region of 5-HT2A, which is predicted to be unstructured. This region has been shown to interact, in vitro, with PSD95 and leads to differences in signaling (Xia et al., 2003). The ASK motif, a component of the tail region, is also important for regulation
-
Despite the significant increase in Bdnf mRNA
2020-05-09
Despite the significant increase in Bdnf1 mRNA levels after DNMT inhibition, the expression of Bdnf4 and Bdnf9 was unaltered. It is particularly surprising that Bdnf4 is not affected by the DNMT inhibitor, as the promoter of the Bdnf4 exon is known to be susceptible to epigenetic changes (Kotera et
-
As part of a program to explore
2020-05-09
As part of a program to explore the key binding interactions between the core of 1 and DGAT-1 a systematic analysis of the aminopyrimidine subunit was initiated. A potential approach to improving passive permeability within this series would be to increase lipophilicity via incorporation of substitu
-
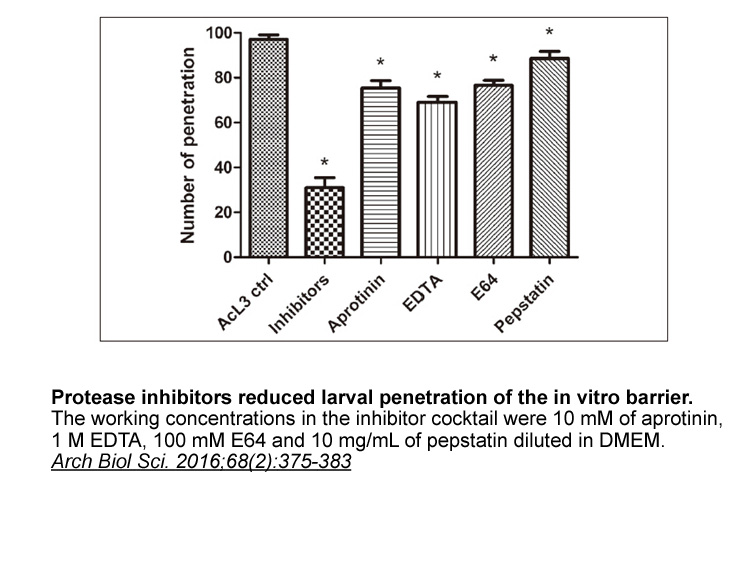
br Materials and methods br Results br Discussion
2020-05-09
Materials and methods Results Discussion GOS potently inhibited SRD5A1 in COS-1 cells (Fig. 4A) and rat Leydig cells (Fig. 4B), possibly attenuating testosterone activation into DHT. It has been reported that gossypol inhibited human SRD5A1 with IC50 of 7 × 10−6 M [28]. Interestingly, gossy
-
Introduction Formate dehydrogenase enzymes FDHs are a group
2020-05-09
Introduction Formate dehydrogenase enzymes (FDHs) are a group of heterogeneous proteins that catalyse the reversible formate () oxidation to carbon dioxide () (Eq. (1)). These enzymes are classified in two families, one gathering the enzymes that hold no redox-active centres and another that includ
-
br Results and Discussion br
2020-05-09
Results and Discussion Materials and Methods Acknowledgements Introduction Inorganic pyrophosphate is generated as a by-product in several enzymatic reactions in cells. These reactions include DNA and RNA synthesis, activation of fatty acids and amino acids, as well as synthesis of the c
-
Plants Cp are widely distributed
2020-05-09
Plants Cp are widely distributed in the plant kingdom and are believed to act as virulence/defense factors for both hosts and pathogens. Cp are also found in plants, animals and bacteria and are known to be virulence factors involved in bacterial pathogenicity [29]. Further, Cp are involved in pepti
16146 records 875/1077 page Previous Next First page 上5页 871872873874875 下5页 Last page